端脑
出处:按学科分类—医药、卫生 中山大学出版社《临床人体解剖生理学》第582页(17172字)
(一)端脑的外形
人类的端脑(telencephalon)(或称大脑,cerebrum)高度发达,是人类思维、记忆等的物质基础,位于颅腔内,脑的表面尚有三层膜,故得到很好的保护。
大脑由前后走向长而深的大脑纵裂(图12-44A、B)(cerebral longitudinal fissure)或半球间裂(interhemispnaric fissura)分成左、右大脑半球(cerebral hemisphere),但在中间部有胼胝体(corpus callosum)相连。脑的表面有凹陷的沟(sulcus)或裂(fissure)。沟与沟间的隆起称脑回(gyrus),其中最主要的沟与裂有:中央沟(central sulcus)、外侧沟(lateral(sylvian)fissure,sulcus)、顶枕沟(裂)(parieto-occipital sulcus,fissure)及扣带沟(cingulate sulcus)等,它们是大脑分叶的标志(图12-44~图12-46)。

图12-44 大脑半球(A、B)

图12-45 大脑半球外侧面观

图12-46 大脑半球内侧面观
每侧大脑半球有“三个面”、“三个极”及“六个叶”。
(1)三个面:上外侧面(superolateral surface)、内侧面(medial surface)及底面(basal surface)(图12-44~图12-46)。
(2)三个极:额极(frontal pole),额叶最前端;枕极(occipital pole),枕叶最后端;颞极(temporal pole),颞叶的最前端(图12-45)。
(3)六个叶:
a.额叶(frontal lobe),位于中央沟前方,外侧裂上方,主要的脑沟与脑回如图12-44~图12-46所见有:中央前沟(precentral sulcus),中央前回(prccentral gyrus),额上沟(superior frontal sulcus),额下沟(inferior frontal sulcus)及额上、中、下回(superior,middle and inferior frontal gyrus)等。
b.顶叶(parietal lobe),位于中央沟后方,主要脑沟、脑回如图12-44~图12-46所示有:中央后沟(postcentral sulcus)、顶内沟(intra interparietal sulcus)、中央后回(posterior central gyrus)和缘上回(supramarginal gyrus)、角回(angular gyrus)等。
c.枕叶(occipital lobe)主要位于半球内侧面、顶枕沟之后,如图12-45、图12-46所示。主要沟、回有:距状沟(calcarine sulcus)、楔叶(guneus)及舌回(lingual gyrus)等。
d.颞叶(temporal lobe),位于外侧沟下方,主要沟、回如图12-45、图12-46所示。所见有:颞上沟(superior temporal sulcus)、颞下沟(inferior temporal sulcus)、颞上回(superior temporal gyrus)、颞中回(middle temporal gyrus)及颞下回(inferior temporal gyrus)及枕颞内侧回(medial ocipitotemporal gyrus)、梭状回(fusiform gyrus)。
e.岛叶(insular lobe)(图12-47),位于外侧裂的深处,被额、顶、颞叶所掩盖。

图12-47 脑岛
f.边缘叶(缘叶)(1imbi c lobe)(图12-46),包括扣带回(cingulate gyrus)、海马旁回(parahippocampal gyrus)和海马旁回钩(uncus),围绕胼胝体(corpus callosum),是组成边缘系统(limbus system)的主要部分。
(二)大脑皮质的内部结构
大脑皮质(crebral cortex)是被覆在半球表面的灰质,它们都是多极神经元,具有分层的形态结构特征。人类大脑皮质总重量约600g,约占脑重的40%。大脑皮质总面积约2200mm2,其中约1/3(750mm2)露于表面,约2/3(1450mm2)位于沟、裂的底和壁。皮质厚度各处不完全相同,平均2.5mm,以中央前回最厚,约4.5mm;距状沟处最簿,仅约1.5mm。大脑皮质神经元总数在120~140亿,如果把所有皮质神经元各自的树突和轴突连接起来,估计总长度有300000~4000000km,相当于地球至月球的距离。
大脑皮质神经元通常分为三大类(图12-48):锥体细胞(pyramid cell)、颗粒细胞(granule cell)及梭形细胞(fusiform cells)。

图12-48 大脑皮质各种神经元分布示意图
(1)锥体细胞(pyramid cell)(图12-48、图12-49),是大脑皮质中特有的一种神经细胞,其形态特征是:胞体呈锥形或等腰三角形,核大呈空泡状,尼氏体(Nissl’s granule)明显。自胞体的尖端发出顶树突或称主干树突(apical dentrit)伸向皮质的表面,沿途分出侧支,接近表面分支较多。在胞体的基部发出一些几呈水平走向的树突称基树突(basal dentrites),并发出分支在胞体附近向四周扩展。在树突表面,尤其在它细小分支上有大量的树突侧棘(dendrite spines),是形成突触之处。有研究表明,丰富的侧棘与皮质功能关系密切,并见到侧棘随年龄增长的发育而增加,在智力减退的婴儿“侧棘”发育受抑制。在胞体的底部发出一条带侧支的长或短的轴突(axon),长轴突伸向髓质组成投射纤维或联合纤维。投射纤维自皮质下行至脑干及脊髓各段,故长者可达1m,在长颈鹿则超过1m。轴突侧支通至皮质各层,故锥体细胞是大脑皮质的主要投射(传出)神经元。
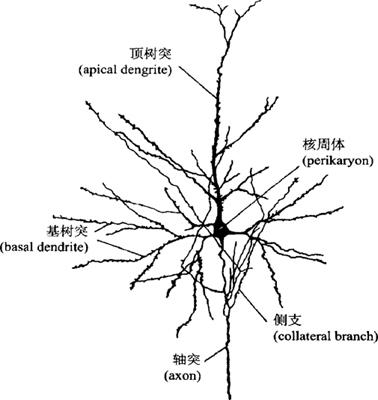
图12-49 大脑皮质的锥体细胞
锥体细胞数量较多,可分为大、中、小三型。大型锥体细胞也称Betz细胞,胞体高100~120μm:中型锥体细胞,胞体高45~50μm;小型锥体细胞,胞体高约10~12μm。锥体细胞数量较多,主要见于皮质IV(锥体细胞)层,也可见于Ⅱ(外颗粒)层和V(节细胞)层。
(2)颗粒细胞(granule cells),数目最多,包括:星形细胞(stellate cells)、篮状细胞(basket cell)、水平细胞(horizontal cell or cajal cell)及Martinotti细胞(图12-48、图12-50),它们共同特征是:胞体较小,直径约6~8μm,呈颗粒状。这类细胞以星形细胞最多,它们轴突多数较短,终止于邻近的锥体细胞或梭形细胞;但有些星形细胞的轴突较长,向上行走向皮质表面与锥体细胞的顶树突或水平细胞相联系。来自丘脑的特异性投射纤维也与颗粒细胞形成突触,所以颗粒细胞是皮质区的中间神经元,构成皮质内信息传递的复杂的微环路。由于人类颗粒细胞发育最盛,是代表皮质种系发生更为进化的结构,也是人类皮质的特征,与人的思维活动密切相关。

图12-50 大脑皮质内各种神经元及相关纤维联系示意图
(3)梭形细胞(fusiform cells)也称多形细胞(polymorphous cells)(图12-48、图12-50),主要位于皮质VI(多形细胞)层。胞体呈梭形,树突自胞体上、下两端分出,上端树突可达皮质表层,轴突由下端树突的主干发出,进入髓质组成投射纤维、联络纤维和连合纤维。
如图12-48、图12-50、图12-51所示,大脑皮质自表到深可依次分为六层:Ⅰ层:分子层(molecular layer),此层神经细胞少,主要是水平细胞和星形细胞、梭形细胞、Martinotti细胞以及皮质一些传人纤维,它们彼此合成较密的纤维,故也称丛状层(plexiform layer)。Ⅱ层:外颗粒层(external granular layer),此层主要由许多密集的星形细胞和小锥体细胞组成。Ⅲ层:外锥体细胞层(external pyramidal layer),也称锥体细胞层(pyramidal layer),此层主要由中小型的锥体细胞和星形细胞组成。Ⅳ层:内颗粒层(internal granular layer),此层较薄,约占皮质全层的10%,主要由星形细胞和篮状细胞组成。从丘脑来的特异性投射纤维在此层水平分支形成致密的横行纤维丛,称Baillarger外带,并与颗粒细胞形成突触。V层:内锥体细胞层(internal pyramid layer)或节细胞层(anglionic layer),此层主要由大、中型锥体细胞以及少量星形细胞和Martinotti细胞组成。Betz细胞也在此层。Ⅵ层:多形细胞层(multiform layer),此层梭形细胞较多,故也称梭形细胞层(fusiform layer),此外尚有星形细胞和Martinotti细胞。

图12-51 大脑皮质的六层结构示意图
(三)大脑皮质的功能区
主要是依据细胞排列类型以及纤维分布形式等的差异作为分区的依据,许多学者如Camphell(1905)将皮质分为20区,Brodmann(1909)分为52区,Economo(1929)分为109区等等,有较大的差异。但以Brodmann的分区应用最广(图12-52A、B)。

图12-52 大脑皮质主要功能区示意图(A、B)
1.躯体运动区(motor area)(图12-52) 主要在额叶。第一躯体运动区或主要运动区(primary motor area)及运动前区(premotor area),位于中央前回即4、6区。躯体各部在运动区皮质的机能定位见图12-53。其机能定位宛如“倒置人体”,即上下颠倒,脚在上,手在下,但面部机能投影位置则依然是正位的;而且是左右交叉,对侧管理,即右侧半球管理左侧半身体运动,左侧半球管理右侧半身体运动;投影区大小与运动的复杂程度成正比。

图12-53 大脑皮质体表运动区定位分布示意图
额叶除躯体运动和语言运动功能外,尚有高级思维功能,它与人的抽象思维和高级智力活动有关。
2.躯体感觉区(somesthetic area)(图12-52、图12-54) 躯体感觉区主要在顶叶的中央后回即3、1、2区。每侧半球接受对侧半身体的本体感觉(位置觉和振动觉等)痛、温觉等,这些感觉在皮质的机能定位,亦如运动区”倒置人体”,但头面部依然是正位的,投影区的大小与感觉的敏感程度成正比。

图12-54 大脑皮质体表感觉区定位分布示意图
顶叶除有躯体感觉功能外,尚与语言等功能有关,尤其在优势侧半球更为明显;也与感觉的整合有关,例如分辨精细触压觉、肢体运动方向及空间定位等。
3.视觉中枢(visual area)(图12-52A、B) 主要在枕叶。第一视觉区或主要视皮质也称纹区(primary visual area,striate area),即17区,位于距状沟两旁,是视觉最主要的皮质区。人识别物象的立体结构能力、识别物体的形状和物体不同部分明暗等,均有赖于第一视觉区。每一侧第一视觉区(纹区)与两眼同侧半的视网膜有联系。实验与临床研究均认为视网膜各部在纹区有一定的投影位置,视网膜上半的纤维止于距状裂的下唇,视网膜下半部的纤维止于距状裂的上唇,而黄斑部的纤维则止于纹状区的后部。第二视觉区或纹旁区(secondary visual area,parastriate area),即18区,它位于17区的上、下方。第Ⅲ视觉区或纹周区(third visual area,peristriate area),即19区,与18区紧邻。目前认为第二、三视觉区对感知和整合视觉是重要的,故被称为视觉联络区。
4.听觉中区(auditory area) 主要位于颞叶的Heschl颞横回,即41区和42区。其中41区是听觉主要的接受区,42区则是听觉的主要联络区,22区紧邻41区,也属听觉联络区(图12-52A)。
颞叶除听觉功能外,尚与语言机能(如Wernicke区)、知识和记忆机能等有关。人类从生活、学习中得来的经验记录,涉及整个大脑皮质的各个部位进行处理,然后被编码存档。目前认为存档部位就在颞叶。
5.与语言运动有关的皮质区 主要位于优势侧半球,即惯用右手的人(称右利者),优势侧半球在左侧;惯用左手,优势侧则在右侧。
(1)运动性语言中枢(motor speech area):位于额下回后部,又称Broca区(44、45区)(图12-52A、图12-55A)。据临床资料Broca区受侵时会发生运动性失语症,即患者虽能发音,却不能说出具有意义的语言。其症状特点是:患者能理解他人的语言,但不能用完整的语言与人对话。一般认为病灶愈大愈易产生失语,失语也愈持久,大多数患者为不完全运动性失语症,病者尚可保存最熟悉的一两个单洞,例如:“好”、“坐”、“不”、“吃”、”再见”等等,但无论如何不能从语音上构成词句。

图12-55 大脑皮质与语言有关的代表区示意图(A、B)
(2)听觉性语言中枢(auditory speech area)(图12-55A、B):位于颞叶及顶叶,包括角回、缘上回及颞上回后部(22、39、40区),也称Wernicke区,它是听觉重要的联络区。据临床观察Wernicke区的被破坏所产生的感觉性失语症比Broca区要严重得多。患者听觉正常,但不能听懂别人和自己说话的意思,故不能正确回答问题和正常说话。“答非所问”是感觉性失语症患者的恰当比喻。
(3)视觉性语言中枢(visual speech area)(图12-55A、B):主要在角回(39区)及其邻近的皮质,它适位于顶、颞叶、枕叶的交界地区,这一区的主要功能被认为是对视觉、听觉语言信息进行整合,从而对语言的意义以及可以表达这些言语的符号进行句法编码,并有目的加以表达。当此区被损害时,视觉没有障碍,但不理解字意、句意。视觉性的语言符号变成毫无意义。例如:患者原来认识的却无法辨认“兄弟的父亲”和“父亲的兄弟”;或看着一幅母子两人的照片,请患者指出“儿子的母亲”和“母亲的儿子”,都会束手无策。临床上常称视性失语症。
(4)书写中枢(writing area)(视性运动语言中枢)(图12-55A、B):位于额中回后部,相当于8、9区。书写是一种复杂的运用功能,书写语言就是写出和声音相当的语言符号(文字),再按通过学习、记忆言语生理过程所安排的次序结合的词和句。所以此区损坏后,患者虽然手的运动机能仍然存在,但写字、绘图等精细动作发生障碍,虽可听懂别人的话和看明白,但不能用写的方式表达,或写出来的字杂乱无章,毫无意义。
人类的语言是大脑皮质整体和长期进化的产物,它是非常复杂的生理过程,而不能认定仅仅是上面论述的几平方毫米的大脑皮质所能完成的。犹如图12-55A、B所示:Broca区不但与唇、舌和咽的运动皮质密切联系,也与Wernicke区密切相关,并与额叶的高级智力活动,与观念形成有关的中枢,以及书写中枢、视性中枢也保持联系。由于人类在各种语言形成过程中依个体的发展而逐渐形成,从幼儿开始先重复其听见的字、看见的字以及幼稚园开始书写的字,然后把些字和某些概念联系起来,当这些观念出现时可用声音、字元,有目的、有条理地表达出来。所以,当与语言相关的皮质有病变时,都会有语言障碍的表现。特别是当上述的几个语言中枢遇到损害,所表现的语言障碍更加明显。
(四)大脑半球的髓质
位于大脑皮质的深面,主要由三种纤维组成:联络纤维、连合纤维和投射纤维。
(1)联络纤维(联合纤维)(association fiber),系指联络本侧半球内不同脑叶、脑回间的纤维(图12-56A、B),例如钩束(uncinate fasciculus)和上、下纵束(superior and inferior longitudinal fasciculus)等。

图12-56 大脑半球内(内侧)联络纤维示意图(A、B)
(2)连合纤维(交连纤维)(commissural fibers),指连合两侧半球间的大纤维束(图12-57)。包括:胼胝体(corpus callosum)是连合左、右大脑半球新皮质的纤维构成;穹隆(fornix)是嗅脑的连合纤维;前连合(前交连,anterior commissure),连接两侧颞叶及左、右嗅球。

图12-57 两侧大脑半球内连合纤维示意图
胼抵体(corpus carosum)(图12-57A、B)是两侧半球间主要的联络通路,估计约有3亿条有髓纤维,被认为对已学习过知识的辨别、感觉经验与记忆有密切关系。主要表现是四肢肌,尤其是手肌技巧动作的协调。所以胼胝体的损害,手的“失用”(apraxic)方面也最为明显。所谓“失用”系指人类在成长过程中所学会的日常生活和生产活动所必需的动作,在长期的反复活动而成为习惯性,并达到显着的自动化程度;以及在一些复杂的动作中运动器官所发生的负反馈的影响,对习惯性动作达到恰到好处的自动调节。其中胼胝体联系两侧大脑半球起着重要的作用,可见如图12-58所示当胼胝体(corpus callosum)受侵害时,患者在试图做出本来习惯有目的或细巧的动作,表现得无能为力、笨拙和动作不连贯,例如让患者用火柴点烟放在嘴里,却用香烟当火柴等等。失用患者不复记得如何应用协调运动作出本来已有观念性动作,即使是最简单的日常动作,也把动作的顺序、时间及动作的组合弄错。所以当出现“失用”并伴有两侧偏瘫,而又不伴有反射增强或Babinski反射时,则应考虑胼胝体受损害的可能。

图12-58 胼胝体病变所引起“失用”示意图
病变在胼胝体(corpus callosum),两侧半球连合纤维中断右半球的缘上回及其邻近皮质失去左半球的影响,引起左侧失用症。
(3)投射纤维(prejection fibers):从广泛的意义上来说应包括由丘脑向皮质投射(传入)的各种感觉纤维以及皮质投射(传出)至皮质下诸结构,如丘脑、纹状体、脊髓等的纤维,但通常指的是:由丘脑各核群直接或间接与大恼皮质发生联系,称为丘脑皮质投射。根据丘脑各部分向大脑皮质投射特征的不同,可分为特异性投射和非特异性投射两大系统。但两者机能上是密切联系的。特异性投射系统的形态学特点是:它们投射至大脑皮质的特定区域,主要是丘脑与大脑中央后回之间,具有点对点的投射关系。非特异性投射系统由脊髓上行的特异性感觉纤维经过脑干时发出与脑网状结构发生联系,经过多次交换神经元后至丘脑中线核群。由丘脑的中线核群,最后弥散地投射到人脑皮质的广泛区域,它们与大脑皮质之间不具有“点对点”的特征关系。
a.特异性投射(传入)纤维:包括浅感觉投射纤维(痛觉、温度觉、粗触压觉),见图12-59;深感觉(本体)投射纤维(来自肌、肌腱关节)等的识别性触觉(tactile discrimination)、两点识别觉(two-point discrimination)、立体感觉(sterognosls)、位置觉(position sense)等,见图12-60、图12-61。

图12-59 浅部感觉传导通路(脊髓丘系和三叉丘系)

图12-60 深部感觉传导通路示意图(内侧丘系)

图12-61 头、面部浅、深感觉传导示意图
来自皮肤的痛、温觉纤维进入脊髓后角(Ⅰ、V、Ⅶ、Ⅷ层)交换神经元后,越过白质前连合,交叉至对侧,形成脊髓丘脑束(spinothalamic tract),上行至丘脑的腹后外侧核,交换神经元后纤维经内囊后肢,投射至中央后回。在脊髓侧索,来自身体下部的浅感觉纤维排列在外侧,上部的纤维在内侧,由内至外按颈、胸、腰、骶有序排列,形成所谓末交叉的长纤维离心性排列(图12-59、图12-62)。

图12-62 浅感觉传导路中在脊髓交叉后的长纤维离心性排列示意图
来自肌、肌腱相关节等的深感觉纤维(薄束、楔束)在延髓下部薄束核、楔束核交换神经元后,越过中线至对侧形成丘系交叉(decussation of medial lemniscus),交叉后的纤维形成内侧丘系(medial lemniscus),上行至丘脑腹后外侧核交换神经元后,经内囊后肢投射至中央后回和中央前回。第一级感觉纤维在脊髓后索的排列是:自身体下部来的深感觉纤维排列在内侧,上部的纤维在外侧,由内向外按骶、胸、颈有序地进入脊髓后索,形成所谓未交叉的长纤维向心性排列(图12-60、图12-63)

图12-63 深感觉传导路中在脊髓交叉后的长纤维向心性排列示意图
b.从浅感觉(痛、温觉)经白质前连合纤维离心性排列及深感觉(本体觉)传入纤维“向心性”在脊髓内排列的规律性,在鉴别脊髓内或外的病变有实用意义。髓内病变,如脊髓空洞症(syringomyelia),系由于先天性发育异常所引起的慢性进行性脊髓疾病。病变初期仅侵犯在前连合,阻断了在此交叉的“痛、温觉纤维”,但触压觉并不受影响,形成所谓节段性分离性感觉障碍(segmental of dissociated sensory disturbance),这是脊髓空洞症最具特征性的临床体征。当病变进一步发展影响到侧索时,感觉痛、温觉丧失常常是自上往下逐渐扩大。脊髓外的病变如硬膜内脊髓外的肿瘤(tumor),若肿瘤位于脊髓后方时可出现后索受损而首先出现精细触觉、意识性感觉相位置觉等深感觉障碍,而痛、温觉保存,即所谓“后索型感觉分离”(posterior funiculus of spinal cord of sensory isotation)。其深感觉障碍出现的顺序是自下而上逐渐扩展。为了便于记忆和应用,综合图12-62、图12-63及上述感觉传导纤维在脊髓内交叉或不交叉的排列情况,可概括为:感觉传导,三级接力,二次交换,一次交叉,全经丘脑,中央后回,对侧管理。若能对照图文理解其意,必能灵活应用。
角膜反射(corneal reflex),是临床上检查昏迷患者深度的方法之一。它由三叉神经、面神经共同完成(图12-64),如果角膜反射消失,说明昏迷程度较深。

图12-64 角膜反射示意图
c.自大脑皮质投射(传出)至皮质下构成的纤维束,有锥体系和锥体外系(pyramidal system and extrapyramidal system)。如皮质至丘脑、纹状体、脑干、脊髓等,其中锥体系(pyramidal system)最为重要。
i.锥体系(pyramidal system):人类一切有目的的随意运动,都是由大脑通过锥体系的传导至所支配的肌肉来完成,所有的哺乳类都是如此,尤其在人类,这种随意运动占绝对优势,所以它对人类高度技巧性动作极其重要。主要起自中央前回大脑皮质到脑干、脊髓运动神经元的最为重要的传导束,包括皮质脊髓束(corticospinal tract)、皮脑脑干束(corticobulbar tract)或皮质核束(corticonuclear tract),见图12-65。

图12-65 锥体系
从锥体系的形态学结构以及临床应用上,对锥体系应明确三点:
(i)其传导径路可概括为:运动传导,二级接力,始自前回,下经内囊,一次交叉,一次交换,精细动作,对侧管理。若能对照图文细解其意,必能灵活应用于临床。
(ii)锥体系全程有两个神经元,即上运动神经元(皮质神经元或中枢神经元)(upper motor neuron)及下运动神经元(或初级运动神经元)(inferior motor neuron),对随意运动的控制,特别是对手技等功能动作的调控,具有非常重要的作用。
上运动神经元是指起自锥体细胞及其下行轴突至下运动神经元之间的结构,如皮质脊髓侧束和皮质脑干束自锥体细胞轴突下行组成锥体束,经内囊、大脑脚等直到脊髓的前角,所以在这些结构中任何一个解剖位置被损害,都会出现上运动神经损害的体征。
下运动神经元是指脊髓前角运动细胞和脑神经运动核及其发出的脑神经和脊神经运动纤维。脊髓前角运动神经元是所有运动传导通路的下运动神经元,是脊髓内各种反射弧(不包括内脏传出)的最后一个环节,即“最后公路”(final common pathway)。这部分损伤可出现下运动神经元损害的体征(图12-66)。

图12-66 传出通路的下运动神经元“最后公路”示意图
(iii)上、下运动神经元损害后的体征截然不同。其要点是:
上运动神经元:损害的部位可以是皮质运动区或锥体束,故瘫痪范围较广泛,表现为随意运动丧失,肌张力增高,为痉挛性瘫(硬瘫);腱反射亢进,浅反射减弱或消失,有病理反射(Babinski征阳性);肌肉与营养障碍而不萎缩,或肌萎缩出现较晚。
下运动神经元:损害部位可以是脑神经运动核或前角运动细胞及相关的神经,故瘫痪范围常较局限,由于肌肉失去了冲动,表现为肌张力减低,为弛缓性瘫痪;腱反射减弱或消失,病理反射(Babinski征阴性);肌萎缩出现早。
在此应特别注意的是,Ⅲ~Ⅻ脑神经运动核中仅面神经核下半部和舌下神经核接受对侧(单侧)皮质脑干束的联系,其他脑神经运动核则接受双侧皮质脑干束的支配,这一点对鉴别核性和核上性损害有重要的临床意义。例如:当患者出现面下部瘫痪,即病为对侧面下部肌瘫痪,鼻唇沟变浅,口角下垂,张嘴无力,但眼裂以上面肌完好,闭眼、额纹、皱眉尚好;令患者伸舌时,舌尖偏向患侧,有时可出现肢体偏瘫和偏身感觉障碍,则病变定位在内囊(核上性)基本可确定。如果核性或核下性侵害则面肌都表现瘫痪,提示病变定位不在内囊(图12-67A、B)。
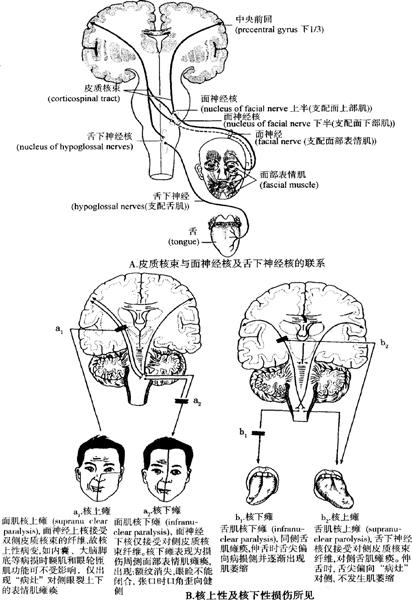
图12-67 皮质核束与面神经核、舌下神经核联系情况及核上性、核下性损伤特点示意图(A、B)
ii.锥体外系(extrapyramidal system):是指锥体系以外的与躯体运动有关的传出通路,在种系发生上,它是较古老的结构,始见于鱼类,到了鸟类是控制全身运动的主要系统;直到哺乳类,尤是人类由于大脑皮质和锥体系统高度发达,锥体外系才逐渐处于从属的地位。在人类锥体外系主要的功能是调整肌张力,协调肌肉活动,维持人体正常姿势和习惯动作。例如走路时下垂双手协调的自然摆动等。但锥体外系与锥体系是功能上密切相关不可分割的一个整体,只有在两者紧密协调配合的情况下才能完成人类日常生活中各种精确的随意运动,例如写字、刺绣、穿针引线、骑自行车、游泳等。其形态学特点是:起于大脑皮质的广泛区域,涉及脑内许多结构,主要有纹状体-苍白球系、皮质-脑桥-小脑系、脑干的网状结构等,以及它们的纤维联系(图12-68)。

图12-68 锥体外系:皮质-脑桥-小脑示意图
(4)内囊(internal capsule):系指位于丘脑、尾状核和豆状核之间进、出脑的白质纤维,如图12-69所示,在脑的切面上呈横置的”><”字型,尖端指向内侧,前外方者为前肢(脚)(anterior limb),位于豆状核与尾状核头之间,其中有额桥束及丘脑背内侧核和前核分别与前额叶皮质和扣带回相联系的往返纤维(丘脑前幅射)。后外方者为内囊后肢(脚)(posterior limb),位于豆状核与丘脑之间,后脚稍长,按其位置可再分为:豆核丘脑部,通过此部的有自丘脑向顶叶中央后回投射的纤维和皮质脊髓束;豆核后部及豆核下部,通过的纤维有丘脑后放线,至枕叶的视觉纤维、顶枕桥纤维以及枕颞桥束和听放射至颞叶;前、后肢之间为内囊膝部(genu),其中有皮质脑干束。

图12-69 脑水平切面,示内囊的分部及主要纤维束的位置
由此可见内囊形态学的特点是:范围狭窄,各种传导纤维高度聚集紧凑:受病变侵害时引起单瘫极为罕见。由于脑血管疾病所致的脑出血病变常易损害的是内囊膝部与后肢,从内囊传导束的纤维排列,认识临床常出现三偏综合征,即病变对侧偏瘫、偏身感觉障碍和同侧偏盲必然会印象深刻。
(五)基底核
基底神经核(basal nuclei) 也称基底神经节(basal ganglia),位于半球内,成对,左、右各一,它们是位于半球内的灰质块,包括尾状核(caudate nucleus)、豆状核(lentiform nucleus)、杏仁核(体)(amygdaloid nucleus,body)及屏状核(claustrum)(图12-70)。

图12-70 基底神经核
1.尾状核状似弯曲向前的“牛尾”(tail of ox),全长与侧脑室的前角、中央部和下角伴行。前端的膨大部称“头”(head of caudate nucleus),相当于内囊膝部移行为“尾状核体”(body of caudate nucleus),超过丘脑后端外侧明显变细称为“尾”(tail of caudate nuclcus),深入颞叶,组成侧脑室下角的上壁,向前几乎与杏仁核相接。
2.豆状核(lentiform nucleus) 位于岛叶深方,略呈楔形,较宽的基底微凸,朝向外侧,尖指向内侧。豆状核的切面上可见有明显的白质髓板将其分成较大的外侧部称壳(putamen),及呈三角样的苍白球(globus pallidus)。
3.纹状体(corpus striatum) 系尾状核与豆状核的合称,从种系发生上又可分为新纹状体和旧纹状体,二者功能不同,其关系如下:

4.纹状体的纤维联系 新纹状体的传入纤维,主要来自大脑皮质、丘脑和黑质,如皮质-纹状体纤维、丘脑纹状体纤维及黑质纹状体纤维等。传出纤维主要是通过纹状体-苍白球纤维、纹状体-黑质纤维与苍白球、黑质相联系。苍白球的传入纤维主要接受新纹状体的传出纤维,苍白球的传出纤维主要有苍白球丘脑纤维、苍白球被盖纤维、苍白球丘脑纤维以及苍白球黑质纤维等。
5.基底核损伤患者常分为新、旧纹状体损伤两种情况:
(1)新纹状体病变表现为运动过多张力障碍综合征(hypokinetic syndromes),如舞蹈病(chored)。
(2)旧纹状体病变表现为运动低下强直综合证(hypokineticrigid sydromes),主要是肢体震颤,肌力升高,运动徐缓及运动不能,尤其是手部及手指,如Parkinson病。
Parkinson病的病变主要位于对侧的黑质(substantia nigra),但也伴有苍白球的损伤。神经化学研究显示,Parkinson病患者的黑质细胞减少(但目前尚未查明原因),引起合成多巴胺(dopamine)的能力减退,以致黑质和纹状体中多巴胺含量均明显降低或消失,这也许是Parkinson病的基础。临床上用多巴胺的前体左旋多巴(levodapa)治疗该病有明显的疗效,其理论基础也在于此。在正常情况下,黑质和纹状体所含的多巴胺,占脑内总量的80%以上。并认为纹状体中的多巴胺来自黑质的大细胞,并由黑质-纹状体纤维运送至尾状核和壳核,并储存于终末的膨体内。
(六)边缘系统
(1)边缘系统(limbic system),主要由边绿叶(limbic lobe),以及与边绿叶皮质结构相似的区域,且又有功能联系上较密切的一些皮质下结构统一的功能系统组成。边缘叶位于半球内侧面,是一相当恒定弯曲的脑回,如图12-71所示,包括:扣带回(cinguli gyrus)、海马旁回(parahippocampal gyrus)或海马回(hippocampal gyrus)、海马钩(uncus)及海马结构(hippocampal formation)等连接成穹隆状的脑回,因其适位于大脑与间脑交界处的边缘,故称边缘叶。

图12-71 边缘系统
(2)属于边缘叶皮质下结构有:额叶眶部、岛叶、颞极、杏仁核簇、隔核、下丘脑、上丘脑、丘脑前核、中脑被盖内侧区等。由于边缘系统在种系发生上比较古老,在功能联系上又十分复杂,它不仅与嗅觉有关,也与内脏活动有关,故它有嗅脑或内脏脑之称。目前还发现边缘系统与情绪行为活动和记忆等也有密切关系。
(3)海马结构(hippocampal formation)(海马体)由海马(hippocampus or ammonishorn)、齿状回(dentate gyrus)以及下托(脚)复体共同组成(图12-72)。

图12-72 海马结构示意图
(由外向内可见海马、海马伞、齿状回、海马沟和海马旁回)
a.下托(脚)(subiculum):是指位于海马旁回皮质和海马之间过渡地区。海马旁回新皮质,有六层结构,但海马和齿状回属旧皮质,仅有三层。而下托(脚)则正好位于此二者之间的移行区。因此,按其移行变化情况将下托(脚)再分为四个带,即旁下托(副下脚)(parasubicum)、前下托(脚)(presubiculum)、下托(脚)(subiculum)和下托尖(最下脚)(prosubiculum)。海马皮质结构具有三个基本层细胞,即丛状层(plexiform layer),位于脑室膜的深面与室床(槽)(alves)相邻,由锥体细胞轴突及传入纤维构成;锥体层(pyramidal layer)由锥体细胞构成;分子层(stratum moleculare)。
海马(hippocampus)又称Ammon,是构成海马结构的主要部分,也是形成侧脑室下角底部最显着的结构。它自胼胝体压部伸至侧脑室下角前端,在此并与海马钩的脑室面相连接。海马的脑室面覆有室管膜,膜的深面是一层白质称室床(槽)(alveus)。室床的纤维向后内方聚集,形成纵行的海马伞,向后汇集成穹隆脚(crus of fornix)。
b.海马结构的纤维联系:传入纤维主要来自内嗅区(entorhinal area),这是最主要的(图12-73),此外还有来自隔内侧核(medial scptal nucleus)、海马的纤维等;传出纤维传统上认为穹隆是海马主要的传出纤维,除终止于乳头体外,尚有纤维止于扣带回、隔核、丘脑前核等。

图12-73 海马下托(脚)划分示意图
边缘系统(limbic system)由于其联系和结构很复杂,对这些不同结构与边缘系统所表现的各种功能之间的相互关系,目前尚未十分了解。从刺激边缘系统的不同部位,可引起呼吸的改变、以及胃肠运动和分泌的改变等。这些内脏反应的表现,主要是通过下丘脑调控。动物实验表明刺激边缘系统的某些部位或切除某些部位,可见动物行为,如与嗅和进食有关的嗅、舐、咀嚼和吞咽以及也表现出有恐惧、逃避、愤怒与攻击等现象:在脑内埋藏电极实验中,也发现在边缘系统有“好感中枢”或“奖赏中枢”(rewarding centers)、“惩罚中枢”(punishing centers)或“恶感中枢”;当刺激前者时动物表现得十分愉快舒服.如果教他通过按电钮方法自我刺激,它很快学会,并不停按电钮;但当刺激后者时,动物表现为嚎叫发怒、瞳孔散大,并立即关断电钮停止刺激。
“好感中枢”的基本部位在丘脑下部腹内侧、杏仁核簇、隔核等,“惩罚中枢”位于丘脑下部的穹隆周围(perifornical nucleus)和中脑的中央灰质等。从各方向看,边缘系统在维持个体生存和种族延续上是重要的,也与人类的情绪活动、内脏活动以及涉及记忆能力等方面密切相关。