中枢耳蜗-前庭途径
出处:按学科分类—医药、卫生 军事医学科学出版社《耳解剖学手册》第186页(6018字)
自从进入脑干后,耳蜗及前庭途径相互分开。
一、耳蜗途径及其中枢(图11-10、11-11)

图11-10 脑干:耳蜗及前庭核群位置

图11-11 耳蜗中枢束(径)(桥脑内段)
在耳蜗神经干交叉越过前庭神经纤维后分成内前和上二支,耳蜗纤维(脑桥神经元)至延髓-脑桥灰质。继续讨论关于耳蜗核群、耳蜗腹束、耳蜗背束、反射束。
1 .耳蜗核群属于两组
(1)腹核:呈顶在下方的锥体形,位于延髓和脑桥之间,在下小脑脚或绳状体上方的前外侧面后方和背核下方之间散开。在此腹核除外侧前庭核的两束纤维外(Hardy纤维来自球状囊并借入耳蜗神经),接受全部第八对脑神经的耳蜗神经纤维。自其上部发出耳蜗腹束或感觉束。
(2)背核:卵圆形,位于腹核的后方。在下小脑脚下方散开以达到第四脑室底外侧隐窝的后方。位于听结节凸起。仅接受来自腹核的纤维。
这些核群为双极细胞体。
2.耳蜗腹束或主要途径
此为到皮质的感觉束。为叙述方便,可分四部认识:
(1)内延髓-脑桥部:此部集中在脑桥橄榄体处,取名为斜方体,其纤维在此与内侧丘系交叉,可分为(据C.Eyries):①同侧纤维,其一部丘系进入橄榄体,在此形成中转站;另一部直接升入外侧丘系。对侧纤维,多出2.5倍。此部大部分纤维中转于橄榄体。一小部分纤维直接进入对侧外侧丘系。②橄榄体间的纤维连接两橄榄体。
接近腹核,主要耳蜗束交叉及继续的前庭束,于此向后者发出一纤维束(Oort)。此束来自耳蜗并获得前庭神经的纤维。
(2)桥脑-丘脑部:组成外侧丘系。其由以前的同侧和对侧外侧耳蜗纤维组成。这些纤维进入内侧膝状体。
起源:起自延髓-脑桥区,在橄榄顶。①直接在小脑束的内方;②在内侧丘系的外方;③在脑神经核前方;④在脑桥神经核后方。
行程:上升斜向前外方,横过脑桥和脚的帽顶,由外侧之内侧丘系散开,同对侧外侧丘系交换纤维。
在脑桥区域内,位于:上小脑脚前方,斜向上内方到红核,由交叉小脑束将其分开。在黑质的后方,借运动束与所在的小脑脚相邻:在内侧丘系的外方。自行连结成一短路;在脚的外内侧是Reil三角区,界邻:①上边的下丘体臂;②前边的小脑脚外侧沟;③后方的上小脑脚隆起。横断面呈顶在外的三角形。
终止:内、外侧丘系相互分开,互相变成直角而止于下丘部;大部分纤维在下丘臂里随行到内侧膝状体。
内侧膝状体下叶的灰核群含有丘脑-皮质束的细胞体或第3神经元。
(3)丘脑-皮质束:形成听放射;横行穿过丘脑下部,在视放射前方获得内囊带状核下的部分止于颞叶皮质。
(4)听皮质区:此区投影在颞叶第一回的上部分、中脑沟的下缘(Heschl横颞灰质,Brodmann 41区)。
听识别区位于颞叶第一回的前区2/3(22区)。
24区是否邻接下后方之14区,尚有争议。这也影响到识别力。
每个耳蜗的皮质投影区都是优势交叉两侧性的。可确定对各种不同声音的相应皮质区(皮质耳蜗);高音与颞横回的内部相对应,低音与外部相对应。
每个耳蜗所产生的纤维均通向两侧皮质区,一侧听区破坏不会导致明显的耳聋。只有当两侧听区破坏时,才会引起全聋。
3 .耳蜗背束:这是一条直接连接耳蜗系统和网状结构的延髓途径,此“唤醒途径”阻碍腹感觉束。它似乎在立体音听觉中起着很重要的作用。
背核产生的纤维混入前边的下小脑脚和后方之第四脑室底之间以形成听纹(髓纹),然后纤维进入缝内以便加入网状结构。此束严格地保持着一侧性。
4.反射束:此束主要司听觉出发点的反射,可能司耳蜗相互作用反射。此束有3组中枢,在3个不同地方。
(1)上橄榄体:由脑桥橄榄、副橄榄、前橄榄核、斜方体核组成,位于延髓-脑桥区。
其传入纤维来自同侧及对侧的耳蜗腹核;传出纤维分布到脑干及其各传导束路。
脑干中有①面神经的两个核,说明镫骨肌反射是两侧性的;②下达脊髓的前角(借后侧纵束);③延髓-桥、眼-脑灰质核;④网状结构中;⑤脑神经植物核中;⑥对侧橄榄体中。
除脑干外,在耳蜗:神经纤维束直接传出耳蜗-橄榄束和Pasmussen交叉束。
(2)丘系核:连接桥脑脚的丘系。其传入纤维来自外侧丘系,传出纤维分布到同侧内侧膝状体的上部和对侧下丘系。
(3)下丘系核群:由主要核、中枢核及其接受外丘系的大量纤维和内-背核组成。其传入纤维来自同侧外丘系和对侧丘系和对侧丘系核,其传出纤维分布到:①对侧下丘:听神经间的Gudden连合。②上丘(听-视束)。③脑神经核及髓质中:桥脑-皮质束、盖-延髓束以及盖-脊髓束。
同侧中脑,特别是肌张力反射的源泉。
二、前庭束及前庭中枢(图11-12,11-13)
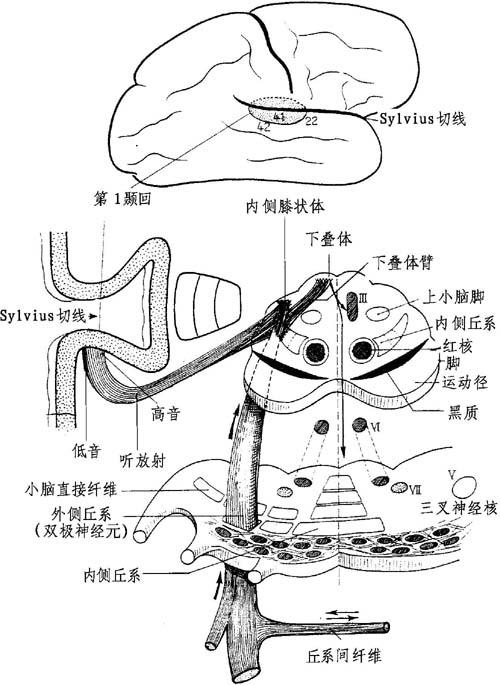
图11-12 耳蜗中枢束(径):脑桥、丘脑及丘脑皮质段
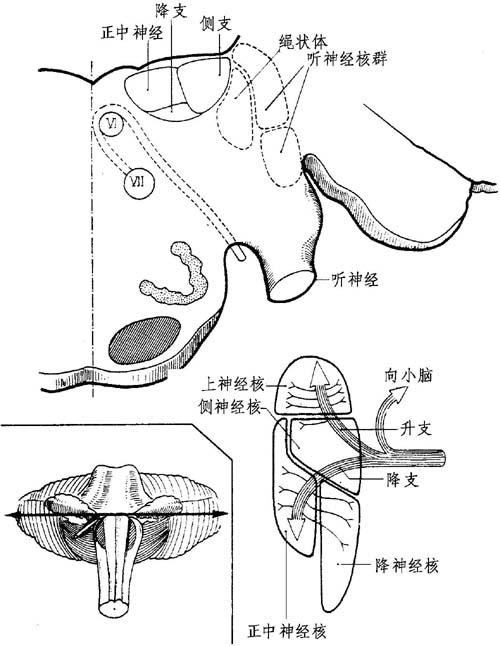
图11-13 前庭神经核
自其进入延髓-脑桥沟后第一级前庭纤维(前神经元)取背部并中间方向进入下小脑脚(绳状体)和三叉神经降根之间。
1 .前庭支:前庭纤维行向前庭外侧核群。
大量纤维束均通向小脑弓,然后自其前庭核群间自行分成升和降两支。这些升降支向同侧发出纤维并横行穿进其所属目标:①到升支的有上前庭核和前庭核的上部分,②到降支的有内前庭核和前庭核下降的部分。
2.前庭核群:这里包括第2级神经元的细胞体(双极神经元)。
(1)位置:这些核形成一个总体或前庭核体群,位于:第四脑室外角和底的下方,在上延髓和下脑桥处。
此等核群与:①上方的三叉神经运动核和上小脑脚;②下方的孤立核和楔状核;③内方外展神经核和面神经膝;④外方的下小脑脚(绳状体);⑤前方的三叉神经降根及感觉核等有关系。
(2)组成:此核群由4组主要核群组成。
①上前庭核(Betcherew),在第四脑室外侧角。大部分出自上核的纤维取道同侧内侧纵束以加入动眼神经核群中。
②内前庭神经核(Schwalbe或三角核)恰好位于第四脑室底下方,在外灰白翼处,对着外展神经核。
此核群发出升纤维在中线交叉以连接对侧纵束,然后到动眼神经核;发出降纤维再形成前庭-脊髓内束。
③前庭降核(Roller),位于内核群外侧,下降得更低。由此出发的纤维到小脑。
④前庭侧核(Deiter)位于前庭核群的外角,在上核和降核之间。
此前庭侧核,实际上不接受前庭神经同侧纤维,也不是真正的第1级前庭核;自此发出前庭脊髓侧索。
⑤下列副簇均连接前庭核群,有在前庭神经之间和f、x、y、z细胞簇之间的前庭神经间质核群。
⑥顶核或盖核,位于前庭区的外侧,在小脑蚓内和小脑橄榄体上方。
此核群同前庭神经终末之间直接相连由此衔接前庭核群。
(3)核间连合的连接:两侧前庭结构是通过:
①一部分对称核间的直接相联关系;
②另一部分,或者网状结构,或小脑的间接关系进行协调。
(4)前庭核群的系统化:前庭神经终末所在局部解剖地位已可在猴和猫身上得到确证,是其每个外周感受野的起始纤维(壶腹和斑):①半规管系统的投影主要在于上和内前庭核群以及小脑;②耳石系统的投影在降核中进行。
三、前庭核群的连接(图11-14,11-15)

图11-14 第1、2前庭神经元系统

图11-15 前庭神经束的总观
前庭双极神经元与感觉性传入纤维存在多发性连接,而且动眼神经可参加控制和调节躯干、颈、四肢以及眼部反射运动的平衡。
1 .脊髓的连接:这里要借止于脊髓前角中之两股纤维束来实现。可控制躯干、四肢以及颈部的肌肉。
(1)前庭-脊髓侧束:侧前庭核生自唯一的同侧纤维,重新发出到脊髓各段;其对伸肌有兴奋作用,而对屈肌则有抑制作用。
(2)束内前庭-脊髓束:来自内前庭核,两侧分布;特别行向颈部脊髓和胸部脊髓高处以供背部和肌肉的运动神经元。
2.小脑连接:这里包括传入和传出途径,与小脑弓(绒球-结节叶)有重要关系。
(1)前庭-小脑连接:有两种:
直接的:由前庭升支产生的纤维组成。后者接受小脑下脚并加入盖核。
间接的:此等纤维在其到达盖核前连接降前庭核群。
(2)小脑-前庭连接:产生于小脑弓皮质并加入前庭核群,借2条途径:①同侧的直接途径;②间接途径,重新连接盖核之后并加入两侧的前庭核。
在小脑和前庭核之间有一条本体及定向非常准确的密切连接小脑和前庭核的双重通道。
3 .与动眼核群的连接:内侧纵束(后纵束)保证其直接连接。其组成一很广泛的联合束,自上方的后白连合到下方的颈髓。
前庭核与动眼核的连接是直接由前庭-中脑束(内侧纵束)来保障和交叉的。分布到:①动眼核群;②位于两脑邻近的Darkschwitsch和Cajal束连接。
系统化:
上前庭核(前庭神经上核)仅分布到同侧动眼神经核且主要管理垂直和旋转功能。
正中核(前庭神经内侧核),为两侧性的,对称分布,管理水平运动。
降前庭核(前庭神经脊核)和侧前庭核(前庭神经外侧核),仅起微小的动眼作用,间接连接属多发神经元的连接。此连接避开内侧纵束并有小脑和网状结构连接。
4.与网状结构的连接:直接借网状结构连接侧前庭核而可影响中枢核的结构。
此网状结构在眼球震颤中起重要作用。
另外,网状结构保证着同交感核、迷核、孤立束核的连接,以阐明出现眩晕的植物神经症状。
5 .与脑皮质的联合:前庭皮质束的存在似难争辩,然而其后的途径尚不十分清楚。此前庭-皮质束只接受同一途径的中枢耳蜗束途径;还有一途径到后侧纵束和视丘。
投影在本体冲动的相等集中点内。
注意:必须阻止耳蜗途径的功能,因它可把声的印象传到大脑皮质,而在前庭途径显然是在皮质下区。
应把前庭系统看成是本体感受器的不同成分之一。
中枢耳蜗束,仅代表极其复杂结构的一个图案,在某些动物身上比人的了解更清楚些。中枢及其多发性神经元并非是简单地反射传达器,而是转变反射。不存在皮质“发射装置”,声音的感受是多发兴奋冲动到达的结果。
与其他感官不同,前庭与丘脑接触极少。